PALÉOETHNOBOTANIQUE EN COURS DE
CHARAVINES - LES BAIGNEURS (ISÈRE)
Karen LUNDSTRÖM-BAUDAIS
CNRS
(Rapport de fouilles, 1983)
Mon travail sur les sédiments du site Néolithique de Charavines "Les Baigneurs" a pris, en raison de la richesse et de la diversité des macrorestes de plantes conservés, plusieurs orientations au fil des années :
1. Tout d'abord, l'identification des essences de bois employées dans les constructions et la détermination de leurs saisons d'abattage ;
2. L'identification des essences sélectionnées pour les outils, les récipients et les ustensiles en bois (voir les différents rapports de fouilles depuis 1980). Ces deux opérations contribuent pour une part à la connaissance du paléoenvironnement, des activités de défrichement et à la technologie du travail du bois au Néolithique ;
3. L'identification des espèces de graines et de fruits récupérées chaque année lors du tamisage systématique des sédiments sur la base terrestre. Les cartes de répartition sont en cours de préparation par le CDPA et s'inscrivent directement dans l'étude spatiale des artefacts et des principaux composants des sédiments archéologiques ;
4. Finalement, le tamisage en laboratoire des sédiments prélevés en blocs nous permet d'isoler les graines fragiles contenues dans les fractions grossières mais que le tamisage massif ne permet pas de conserver et les très nombreuses graines des fractions fines. Cette quatrième approche nous offre la possibilité de compléter très largement l'éventail des graines isolées au tamisage massif. Cette approche plus détaillée nous permet aussi d'affiner nos observations sur le comportement stratigraphique des espèces car ces prélèvements sont moins sujets aux processus de pollution par infiltration d'une couche à une autre, comme cela peut survenir parfois dans les cas des graines récupérées lors du tamisage intégral (Lundström-Baudais, 1982).
Le bloc Y
Le présent rapport concerne principalement
ce dernier aspect de nos travaux. En 1980, grâce aux efforts "héroïques"
de l'équipe de plongeurs, un bloc sédimentaire correspondant
à deux triangles métriques, épais de 50cm et pesant plus
de 600 kg a été extrait du site et transporté à
terre (ce bloc contenait la séquence complète des couches archéologiques).
L'échantillonnage a été effectué en présence
des fouilleurs-plongeurs et de A. Bocquet pour assurer la subdivision de la
séquence archéologique la plus complète possible. Au
même moment, J.-L. Brochier effectuait la prise d'échantillons
pour l'étude sédimentologique (Brochier, 1980). Nous avons pris
six échantillons pour l'étude des macrorestes.
Quatre cents grammes de chaque échantillon ont été tamisés
à l'eau en laboratoire avec des tamis de mailles 8, 4, 2, 1, 0.5 et
0.2mm. La séparation des graines, fruits, balle de céréales,
capsules de lin, etc ... a été faite sous la loupe binoculaire.
La totalité des débris végétaux des fractions
8 à 1mm a été triée alors que seulement une moitié
et le quart des fractions 0.5mm et 0.2mm (le nombre de graines compté
dans chacune de ces deux dernières fractions a été multiplié
par le facteur correctif approprié pour faire en sorte que les chiffres
donnés correspondent à un échantillon de 400g). Pour
l'identification de ces restes, je me suis essentiellement servie de ma collection
comparative de graines modernes complétée par les atlas de graines
de Beijerninck, 1976, Berggren 1981, Bertsch 1941, Katz, Katz et Kipiani 1965
et des catalogues descriptifs des plantes de différents sites archéologiques
comme par exemple ceux de Korber-Grohne 1967, van Zeist 1974 et Frits 1977.
Jusqu'à présent nous avons identifié quelques 81 taxons
parmi les restes de bois et de graines des différents systèmes
de tamisage. Pour présenter les résultats, il est essentiel
d'ordonner les espèces par groupements végétaux (tableau
2). Cette ordination des espèces pose certaines difficultés,
même si l'on emploie des groupements extrêmement larges ; souvent
dans la nature les espèces se rencontrent dans plusieurs groupements
végétaux à la fois. Dans cette figure, les groupements
dans lesquels une espèce est susceptible d'apparaître sont indiqués
par un cercle blanc alors que le cercle noir correspond au groupement dans
lequel la plante est le plus fréquemment observée aujourd'hui.
Par ces groupements on passe progressivement de la végétation
du lac à celle des rives et finalement à celle de la terre ferme
et de l'arrière (espèces rudérales, agrestes, des prairies
et des forêts). Nous complétons ce tableau en donnant une indication
sur l'usage potentiel qui peut être fait des différentes espèces
(alimentaire, médicinal, teinture, sparterie). Les espèces marquées
d'un (*) ont très probablement été collectées.
Les 6 échantillons contenaient quelques 26.904 graines, restes de battage
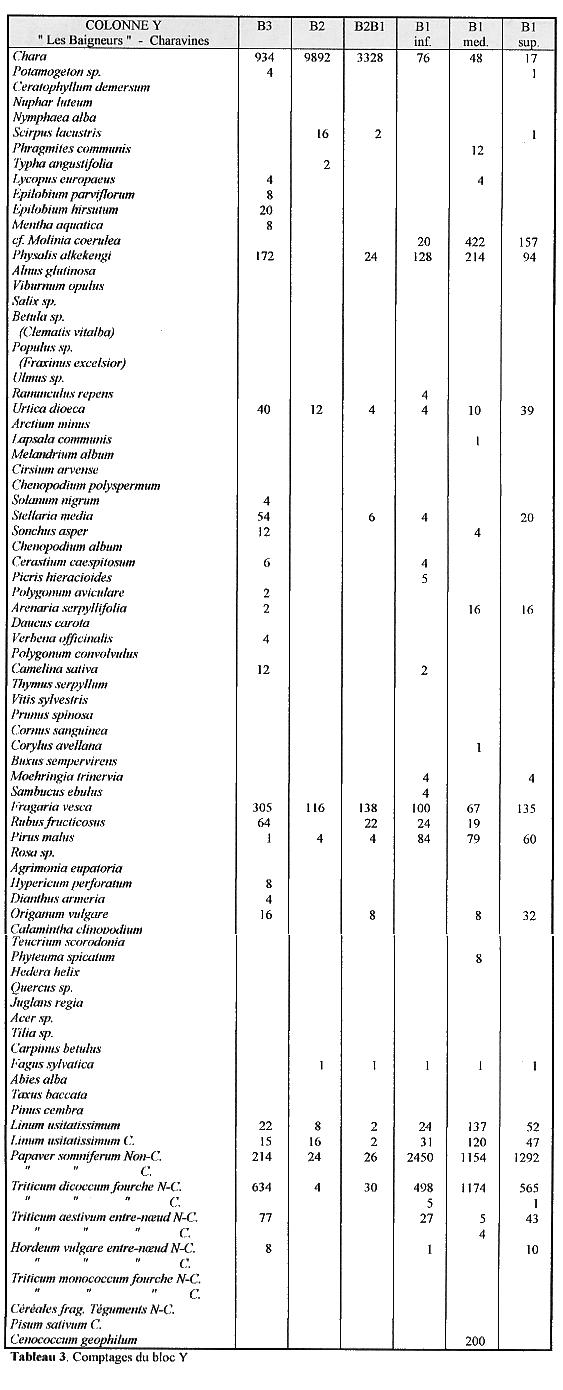
des céréales, oogones et fongi. Les comptages du bloc Y apparaissent
en détail dans le tableau 3.
Nous devons insister sur le fait que les observations que nous présentons ici ne sont que des résultats préliminaires ; dans l'immédiat nous ne disposons que des résultats d'un seul bloc et il est absolument nécessaire d'étendre les analyses pour vérifier et préciser nos premières "impressions".
1. LA VÉGÉTATION AQUATIQUE ET DES RIVES
La végétation aquatique est représentée dans les bloc Y par les oogones de Characées et les graines de Potamogeton sp. Les oogones sont relativement plus nombreuses dans la B3 et dans la partie supérieure de la B1.
La végétation des roselières apparaît avec Scirpus lacustris, Phragmites communis, Typha cf. latifolia. Les traces de cette végétation sont absentes dans la B3. Dans la B2 on observe Typha cf. latifolia et Scirpus lacustris, cette dernière disparaît dans la B1 médiane alors qu'apparaît Phragmites communis. L'interprétation de ces comportements est encore difficile à mener car les informations sont pour le moment trop éparses avec un seul bloc étudié jusqu'à présent. La succession observée ici doit être vérifiée sur d'autres colonnes, toutefois un certain nombre d'observations peuvent déjà être avancées :
- En ce qui concerne les variations dans la représentation des oogones, il est à noter que la diminution ou l'augmentation des Characées n'est pas nécessairement liée à un changement dans le niveau d'eau. Les Characées sont généralement oligotrophes et très sensibles à la turbidité de l'eau. Même sans changements dans le niveau du plan d'eau, l'installation des habitats provoquera leur disparition. Ceci dit, revenons sur l'importance des oogones dans la B3 comparativement à la B1 : les oogones de la B3 sont très certainement une intrusion de macrorestes qui ne représentent pas du tout la végétation sur place. Elles sont peut-être un indice du remaniement des craies sous-jacentes lors de la première installation ou encore après l'abandon du premier village. Dans cette optique, il serait intéressant d'étudier le comportement des macrorestes de la végétation aquatique dans les sédiments qui sont à l'abri des remaniements parce que pris sous les chapes d'argile.
- Les très nombreuses oogones de Characées ainsi que les graines de Scirpus lacustris et Typha cf. latifolia suggèrent qu'à ce moment de son histoire, le site se trouvait plus ou moins en bordure de la zone eulittorale. Ceci se corrèle bien avec les résultats de l'étude sédimentologique, notre échantillon correspond au B2B de l'analyse de Brochier qui suggérait une hauteur d'eau de l'ordre de 1,50m sur le site.
- Dans la B3, l'apparition de Phragmites
communis à la suite de la disparition de Scirpus lacustris peut être
interprétée comme la preuve d'une légère diminution
du niveau d'eau (Scirpus lacustris se trouve généralement au
large de la ceinture de roseaux). Mais on ne peut écarter l'idée
que les graines des roseaux ont été introduites sur le site
par les activités humaines ; par exemple s'ils utilisent les roseaux
comme matériau de construction pour les toitures.
Il est certain que pour établir une image plus claire du comportement
des macrorestes de la végétation aquatique et palustre, nous
devons analyser plus d'échantillons, à la fois sur le site lui-même
et en direction du tombant et de la terre ferme actuelle. Ces échantillons
doivent être pris en bloc ou en colonne pour éviter toute ambiguïté
stratigraphique. Il est également urgent de faire de petits prélèvements
au fond du lac et sur les rives pour observer la répartition des macrorestes
qui se déposent actuellement et leur rapport avec la couverture végétale
qui existe aujourd'hui dans ces zones. Sans modèle de comparaison toute
interprétation risque de rester du domaine de l'hypothèse.
Les déterminations des essences de bois ont clairement montré
que les défrichements ont eu lieu au moins occasionnellement dans la
forêt riveraine. Ces défrichements auront encouragé le
développement des prairies mouillères dont les espèces
suivantes témoignent : Epilobium hirsutum, Epilobium parviflorum, Mentha
aquatica et Molinea coerulea. Cette dernière est très bien représentée
dans les sédiments de la deuxième phase d'occupation du site.
Il est possible qu'il faille mettre l'apparition de cette espèce en
rapport direct avec l'abondance des graminées notée par A. Barbier
dans les pollens des sédiments de la zone tourbeuse à 300m en
retrait du village. Ce phénomène apparaît alors que la
séquence pollinique traduit des activités de défrichement
(voir le rapport 1982) et il a été interprété
comme la preuve de l'utilisation de cette zone riveraine comme lieu de pâturage.
Les graines des plantes aquatiques et de rives représentent 6% de la totalité des graines et restes associés dans les couches archéologiques ; ce pourcentage est accentué par la présence abondante de Molinia coerulea.
Les plantes cultivées comptent pour 7% des graines. Les céréales sont très bien représentées par les restes non-carbonisés de balle (les fourches : l'articulation entre les glumes et le rachis) et les fragments de rachis de l'épi (entre-nœuds : article de rachis). Seulement 0,3% de ces restes ont été carbonisés. Triticum dicoccum, Triticum aestivum et Hordeum vulgare sont les principales céréales et après un rapide survol des céréales carbonisées ramassées au tamisage massif, Triticum monococcum semble n'avoir guère d'importance. Les graines de Linum usitatissimum et Papaver somniferum sont très nombreuses, le lin a très certainement été exploité pour la qualité de ses fibres et pour ses graines. Pisum sativum est absent de l'échantillon Y alors que quelques grains ont été isolés au tamisage systématique. Leur chance de se trouver conservées dépend du degré de carbonisation atteint.
Il est difficile de saisir l'importance de ces plantes alimentaires au travers du seul nombre de graines ou restes associés retrouvé. Pour mieux préciser leur rôle réciproque nous avons converti les restes de balle et les fragments d'épi en nombres de grains auxquels ils correspondent. Ensuite nous avons traduit en poids équivalent ces nombres de grains pour faciliter l'évaluation de l'importance de chaque espèce (tableau 1).
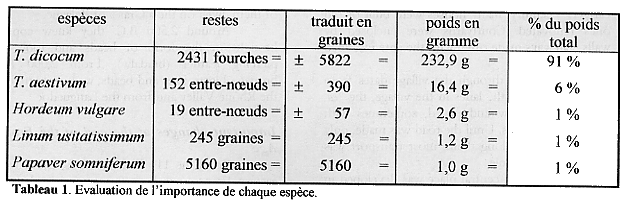
Tableau 1. Evaluation de l'importance de chaque espèce.
Les céréales dominent nettement et sont de toute évidence les récoltes principales. Le lin et le pavot ne sont que des récoltes d'importance très secondaire. Il sera intéressant dans le futur de voir si, avec plus d'analyses, l'orge se maintient dans son rôle relativement insignifiant. Enfin, il s'avérera nécessaire de comparer les résultats obtenus par l'analyse des céréales carbonisées isolées au tamisage systématique de ceux des restes de battage récupérés lors du tamisage fin (notons que dans un kilo de sédiment nous parvenons à isoler l'équivalent d'environ 5.000 grains de céréales sous la forme de restes de battage, ce qui est deux fois supérieur aux nombre de céréales carbonisées isolées lors du tamisage systématique sur l'ensemble du site depuis le début des recherches)
Ces restes de balles et fragments de rachis nous permettent d'affirmer qu'il y a eu stockage et battage des céréales à même le site. Le battage a probablement eu lieu au fur et à mesure des besoins car sous climat humide, les graines stockées dans leurs glumes risquent beaucoup moins de se détériorer que celles qui en sont déjà débarrassées.
L'évidence du stockage et du battage des céréales sur le site a d'importantes implications sur l'interprétation des résultats de l'analyse pollinique. Les forts pourcentages de céréales que l'on observe dans les couches archéologiques sont très certainement liées aux activités de battage sur le site même. Chez les céréales autogames, dont font partie toutes nos céréales, les glumes piègent des quantités importantes de leurs propres pollens. Il y a plus de pollens libérés lors du battage qu'au moment de la pollinisation (voir la discussion Lundström-Baudais 1983). On ne peut donc pas, sur ce site, considérer les pollens comme un indicateur fiable de la distance des champs par rapport au village.
3. LES MAUVAISES HERBES RUDÉRALES
ET AGRESTES
Nous avons rencontré 17 espèces de mauvaises herbes lors de l'expérimentation pour la mise en place du tamisage systématique alors que 10 seulement figurent dans les couches archéologiques du bloc Y, ce qui représente à peine 2% de la totalité des graines.
Ces mauvaises herbes donnent une image qui ressemble peu à celle que nous associons de nos jours aux champs de céréales. Les espèces que l'on retrouve fréquemment dans cette colonne sont aujourd'hui associées aux cultures sarclées et aux jardins, ce sont : Stellaria media, Chenopodium album, Chenopodium polyspermum, Solanum nigrum, Sonchus asper et Verbena officinalis. Seul Polygonum convolvulus est représentatif des cultures céréalières mais cette espèce n'est pas à proprement parler très stricte puisqu'elle se développe aussi sur les terrains sarclés.
La végétation qui se développe sur les sols cultivés est aujourd'hui directement fonction de la date à laquelle la terre a été retournée pour la dernière fois. Sur les champs labourés et semés en automne ce sont les mauvaises herbes, qui germent à relativement basse température, qui se développent (annuelles d'hiver) alors que sur les terrains sarclés et les jardins, où les bouleversements sont tardifs (mai-juin, parfois plus tard), les annuelles d'été dominent largement. Il faut noter que les champs d'hiver qui subissent le sarclage intensif se trouvent dans ce cas. Si le sol est remué tôt au printemps puis laissé en repos - comme c'est le cas des champs de céréales de printemps semés en mars-avril - la flore des mauvaises herbes qui s'y développe est un mélange d'annuelles d'hiver et d'été. Le manque de mauvaises herbes strictement associées aux champs céréaliers d'hiver dans les sédiments des "Baigneurs" semble de toute façon écarter la possibilité d'un ensemencement hivernal qui ne serait pas accompagné d'un sarclage très intensif et efficace.
C'est ici qu'il est intéressant d'introduire nos observations concernant la fréquence des mauvaises herbes dans les sédiments archéologiques. Le nombre de mauvaises herbes dans le bloc Y est extrêmement bas en comparaison d'autres sites néolithiques de la même période que nous avons également analysés (Auvernier "Brise-Lames", NE et Clairvaux station III, Jura). Les restes de plantes cultivées dans le bloc Y sont 25 fois plus nombreux que les mauvaises herbes alors qu'à la station III de Clairvaux ils ne sont que 5 fois plus importants. On peut avancer plusieurs hypothèses pour expliquer cette différence :
- La technique utilisée pour moissonner affecte directement le nombre de mauvaises herbes ramassées avec la récolte. Si c'est uniquement l'épi qui est coupé, l'introduction des mauvaises herbes dans la récolte est très réduite. Si au contraire l'épi est coupé avec la majeure partie du chaume, ou encore, si après la récolte des épis on entre le chaume, le nombre de mauvaises herbes est alors beaucoup plus élevé. Dans le cas de la station III de Clairvaux, nous avons cru déceler une récolte en deux temps, les épis puis le chaume. A première vue, le peu de graines de mauvaises herbes à la station des "Baigneurs" suggère que seul l'épi était récolté. Mais l'importance de mauvaises herbes courtes comme Arenaria serpyllifolia (qui mesure 20cm au maximum) laisse penser que sur ce site aussi le chaume a été ramené au village. Donc la différence dans le nombre de mauvaises herbes entre ces deux sites n'est pas à rapprocher des techniques de moissonnage.
- Les pratiques agraires influencent le nombre de mauvaises herbes qui se développent sur les sols cultivés. L'hypothèse d'une culture sur brûlis a déjà été avancée par A. Barbier sur la base de son analyse pollinique. Sur les champs qui ont subi ce traitement préparatoire, il y a un nettoyage radical des plantes adventices ; le sarclage ou toute autre technique destinée à l'élimination des mauvaises herbes n'est pas nécessaire pour les quatre ou cinq années suivantes.
Donc les récoltes prises sur les champs préparés par brûlis et ensuite exploités sur de courtes durées introduiront avec eux peu de mauvaises herbes sur le site, ce qui est le cas dans les sédiments des "Baigneurs". D'autre part le brûlis provoque une augmentation très temporaire des substances nutritives disponibles qui encouragent une végétation proche de la flore de nos jardins.
Deux procédés agricoles peuvent expliquer la flore des mauvaises herbes de la station des "Baigneurs", soit un sarclage intensif, soit la pratique de la culture sur brûlis.
Il nous est encore difficile de trancher sans plus d'analyses. Bien évidemment, les échantillons pris dans un bloc ne nous donnent pas nécessairement l'image correcte des mauvaises herbes dans l'ensemble des sédiments de ce site.
4. LA VÉGÉTATION DES PRAIRIES
Jusqu'à présent l'étude des macrorestes ne nous indique que la présence des prairies mouillères. L'absence des espèces associées aux prairies plus sèches permet d'envisager le parcage du bétail et le stockage du fourrage (si pratiqué) en dehors du site. La quantité de coprolithes retrouvés pourrait contredire cette hypothèse. Il est plutôt probable que le pâturage se limitait aux prairies mouillères des rives du lac et au sous-bois des forêts avoisinantes.
5. LA FORÊT ET SES LISIÈRES
Si l'étude des bois nous a permis un aperçu des défrichements dans la hêtraie sapinière de l'arrière-pays, l'étude des graines ne divulgue que très peu d'informations. La lisière par contre est très bien représentée dans l'étude des graines car cette végétation produit de nombreuses baies et fruits que l'homme a ramassé pour sa consommation. Les produits de la cueillette comptent pour ou moins 16 % des graines et la majorité des plantes ainsi exploitées provenait de cette zone (tableau 3). De ces 17 espèces seulement cinq n'ont probablement pas été intentionnellement ramassées ; celles-ci ont pu se trouver en bordure des champs et être introduit sur le site au même moment que les différentes récoltes.
6. CONCLUSION
Une fois que nous aurons étudié
un nombre suffisant d'échantillons, il deviendra possible de tenter
l'évaluation de l'importance relative de la cueillette dans l'alimentation
par rapport aux produits de l'agriculture.
Les premiers résultats du tamisage fin font ressortir deux lignes directrices qui doivent être, à notre avis, abordées de manière distincte :
- Premièrement, il y a l'étude du comportement stratigraphique des macrorestes de la végétation aquatique et riveraine qui aborde les problèmes de la mise en place des sédiments et de l'évolution de l'environnement directement aux alentours du site. Pour poursuivre cette étude, il est nécessaire d'étendre le nombre d'échantillons à des emplacements bien sélectionnés. Il faut continuer de travailler avec les échantillons pris en colonne ou bloc pour éviter la contamination entre les niveaux. Le tri, par contre, peut se réduire uniquement à la sélection des macrorestes produits par la végétation de ces zones.
- Les études liées aux plantes cultivées, aux méthodes agricoles ainsi qu'à la cueillette, font l'objet d'un deuxième ensemble. Les refus des tamis fins (1mm et 0,5mm) sont les plus appropriés pour ces études. Ces refus évitent le problème toujours difficile de la représentativité des échantillons ponctiformes et nous permettent d'intégrer nos résultats à l'analyse spatiale du site.
Les observations préliminaires et leurs interprétations ici présentées démontrent bien le potentiel interprétatif contenu dans l'analyse des macrorestes et comment cette étude s'intègre directement aux préoccupations des archéologues qui cherchent à mieux connaître le contexte global de l'homme préhistorique.
BIBLIOGRAPHIE
BROCHIER J.-L., 1980. La station des Baigneurs - Charavines (Isère). Études sédimentologiques. Premiers résultats. In : Bocquet A. (Dir.) : Village néolithique immergé, station des Baigneurs (Charavines, Isère, lac de Paladru). Rapport de fouilles, d'études connexes et d'animations, p. 45-70.
LUNDSTRöM-BAUDAIS K., 1982. Paleoethnobotanique à Charavines-Les Baigneurs. Récolte des éléments et méthodes d'étude. In : Bocquet A. (Dir.) : Village néolithique immergé, station des Baigneurs (Charavines, Isère, lac de Paladru). Rapport de fouilles, d'études connexes et d'animations.
ZEIST (van) W. et CASPARIE W.A., 1974. Niederwil, a palaeobotanical study of a swiss neolithic lake shore settlement., in : Geologie en Mijnbouw, vol. 53, n°6, p. 415-428.